
Forskjell mellom manglende reparasjon og Nucleotide Excision Repair | Mismatch Reparasjon vs Nucleotide Excision Repair
Nøkkelforskjell - Manglende reparasjon vs Nucleotide Excision Repair
Tens og tusenvis av DNA-skader forekommer i cellen per dag. Den induserer endringer i celleprosessene som for eksempel replikasjon, transkripsjon og cellens levedyktighet. I noen tilfeller kan mutasjoner forårsaket av disse DNA-skadene føre til skadelige sykdommer som kreft og aldringsrelaterte syndromer (for eksempel: Progeria). Uavhengig av disse skadene initierer cellen en høyt organisert kaskadreparasjonsmekanisme som kalles DNA-skaderesponser. Flere DNA-reparasjonssystemer er blitt identifisert i cellesystemet; Disse er kjent som Base Excision Repair (BER), Mismatch Repair (MMR), Nucleotide Excision Repair (NER), Double strand pause reparasjon. Nucleotide excision reparasjon er et svært allsidig system som gjenkjenner storskala helixforvrengning DNA lesjoner og fjerner dem. På den annen side erstatter feilparametre feilbaserte baser under replikering. Nøkkeldifferansen mellom feilparametreparasjon og nukleotidutskytningsreparasjon er at nukleotidutskriftsreparasjon (NER) brukes til å fjerne pyrimidindimere dannet ved UV-bestråling og voluminøse helixlesjoner forårsaket av kjemiske addukter mens feilparametere reparerer system spiller en viktig rolle i korrigering av feilinnblandet baser som har rømt fra replikasjonsenzymer (DNA polymerase 1) under postreplikasjon. I tillegg til mismatchede baser kan MMR-systemproteiner også reparere innsatsene / deletionsløkkene (IDL) som er resultater av polymerase-slippingen under replikasjon av repeterende DNA-sekvenser.
INNHOLD
en. Oversikt og nøkkelforskjell
2. Hva er Mismatch Repair
3. Hva er Nucleotide Excision Repair
4. Side ved side-sammenligning - Feilsøking Reparasjon vs Nucleotide Excision Repair
5. Sammendrag
Hva er Nucleotide Excision Repair?
Den mest fremtredende trekk ved nukleotidutjevningsreparasjon er at den reparerer de modifiserte nukleotidskader forårsaket av signifikante forvrengninger i DNA-dobbelthelixen. Det observeres i nesten alle organismer som har blitt undersøkt oppdatert. Uvr A, Uvr B, Uvr C (excinucleases) Uvr D (en helikase) er de mest kjente enzymer involvert i NER som utløser reparasjon av DNA i modellorganismen Ecoli. Uvr ABC multi-underenheter enzymkompleks produserer Uvr A, Uvr B, Uvr C polypeptider.Generene kodet for ovennevnte polypeptider er uvr A, uvr B, uvr C. Uvr A og B enzymer gjenkjenner kollektivt den skaderinducerte forvrengning som forårsakes av DNA-dobbelthelixen, slik som pyrimidin-dimmere på grunn av UV-bestråling. Uvr A er et ATPase enzym og dette er en autokatalytisk reaksjon. Deretter forlater Uvr A DNA mens Uvr BC kompleks (aktiv nuklease) spalter DNA i begge sider av skaden som katalyseres av ATP. Et annet protein kalt Uvr D kodet av uvrD-genet er et helikase II-enzym som vikler DNA som er resultatet av frigjøring av enkeltstrenget skadet DNA-segment. Dette etterlater et gap i DNA-helixen. Etter skadet segment er blitt skåret, forblir et 12-13 nukleotidspalt i DNA-strengen. Dette fylles opp av DNA-polymerase-enzymet I og nikket er forseglet av DNA-ligasen. ATP kreves ved tre trinn av denne reaksjonen. NER-mekanismen kan også identifiseres i pattedyr-lignende mennesker. Hos mennesker er huden tilstanden kalt Xeroderma pigmentosum på grunn av DNA-dimers forårsaket av UV-bestråling. Generene XPA, XPB, XPC, XPD, XPE, XPF og XPG produserer proteiner som erstatter DNA-skade. Proteinene av gener XPA, XPC, XPE, XPF og XPG har nukleaseaktiviteten. På den annen side viser proteinene av XPB og XPD-gener helikaseaktiviteten som analoger til Uvr D i E coli.

Figur 01: Nucleotide Excision Repair
Hva er manglende reparasjon?
Feilsparingsreparasjonssystemet initieres under DNA-syntese. Selv med funksjonell € -underenheten tillater DNA-polymerase III inkorporering av et feil nukleotid for syntese hvert 10 8 basepar. Manglende reparasjonsproteiner gjenkjenner dette nukleotidet, aksepterer det og erstatter det med det riktige nukleotid som er ansvarlig for den endelige grad av nøyaktighet. DNA-metylering er pivotal for MMR-proteiner for å gjenkjenne overordnetstrengen fra den nylig syntetiserte strengen. Metyleringen av adenin (A) nukleotid i et GATC-motiv av en nylig syntetisert streng er litt forsinket. På den annen side har moderstrengadene-nukleotidet i GATC-motivet allerede metylert. MMR-proteiner gjenkjenner den nylig syntetiserte streng ved denne forskjellen fra overordnet streng og starter feilparametre i en nylig syntetisert streng før den blir metylert. MMR-proteiene styrer deres reparasjonsaktivitet for å akseptere feil nukleotid før den nylig replikerte DNA-strengen blir metylert. Enzymer Mut H, Mut L og Mut S kodet av gener mut H, mut L, mut S katalyserer disse reaksjonene i Ecoli. Mut S-protein gjenkjenner syv av åtte mulige mismatchbasepar unntatt C: C, og binder på stedet for mismatch i duplex-DNA. Med bundet ATP, blir Mut L og Mut S med i komplekset senere. Komplekset oversetter noen få tusen basepar bort til den finner et hemimetylert GATC-motiv. Den sovende nukleaseaktiviteten til Mut H-protein aktiveres når den finner et hemimetylert GATC-motiv. Den klipper den ikke-metylerte DNA-strengen, og etterlater et 5'-nick ved G-nukleotid av ummetylert GATC-motiv (nylig syntetisert DNA-streng).Da blir den samme strengen på den andre siden av mismatchen nicked av Mut H. I resten av trinnene aksesserer de kollektive virkningene av Uvr D et helikaseprotein, Mut U, SSB og exonuclease I det ukorrekte nukleotidet i enkeltstrenget DNA. Spaltet som dannes i excisionen fylles opp av DNA-polymerasen III og forsegles ved ligase. Et lignende system kan identifiseres hos mus og mennesker. Mutasjonen av humant hMLH1, hMSH1 og hMSH2 er involvert i arvelig nonpolyposis tykktarmskreft som avregulerer celledeling av kolonceller.
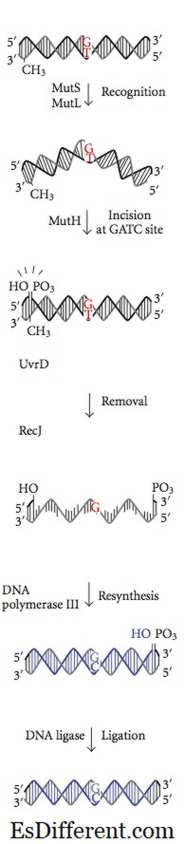
Figur 02: Feilsøkingsreparasjon
Hva er forskjellen mellom Reparasjon av reparasjon og Nucleotide Excision Repair?
- diff Artikkel Midt før tabell ->
Manglende reparasjon vs Nucleotide Excision Repair |
|
| Manglende reparasjonssystem oppstår under post-replikering. | Dette er involvert i fjerning av pyrimidindimerer på grunn av UV-bestråling og andre DNA-lesjoner på grunn av kjemisk addukt. |
| Enzymer | |
| Den katalyseres av Mut S, Mut L, Mut H, Uvr D, SSB og exonuclease I. | Den katalyseres av Uvr A, Uvr B, Uvr C, UvrD enzymer. |
| Metylering | |
| Det er sentralt for å initiere reaksjonen. | DNA-metylering er ikke nødvendig for å starte reaksjonen. |
| Virkning av enzymer | |
| Mut H er en endonuklease. | Uvr B og Uvr C er exonukleaser. |
| Anledning | |
| Dette skjer spesielt under replikering. | Dette skjer når det eksponeres for U.V eller kjemiske mutagenser, ikke under replikering |
| Bevaring | |
| Den er sterkt konservert | Den er ikke høyt konservert. |
| Gap fylling | |
| Det gjøres ved DNA polymerase III. | Det er gjort av DNA-polymerase I. |
Sammendrag - Reparasjon av manglende reparasjon vs Nucleotide Excision Repair
Manglende reparasjon (MMR) og Nucleotide Excision Repair (NER) er to mekanismer som finner sted i cellen for å rette opp DNA-skader og forvrengninger som er forårsaket av ulike agenter. Disse kalles kollektivt som DNA-reparasjonsmekanismer. Nukleotidutskriftsreparasjon reparerer de modifiserte nukleotidskader, typisk de signifikante skader på DNA-dobbelt-helixen som skjer på grunn av eksponering for UV-bestråling og kjemiske addukter. Manglende reparasjonsproteiner gjenkjenner feil nukleotid, aksepterer det og erstatter det med riktig nukleotid. Denne prosessen er ansvarlig for den endelige grad av nøyaktighet under replikering.
Referanse:
1. Cooper, Geoffrey M. "DNA Reparasjon. "The Cell: En Molekylær Tilnærming. Andre utgave. U. S. National Library of Medicine, 01 Jan. 1970. Web. 9. mars 2017.
2. "Mekanismer og funksjoner for DNA-mismatch reparasjon. "Cellforskning. U. S. National Library of Medicine, n. d. Web. 09 Mar. 2017.
Image Courtesy:
1. "Nucleotide Excision Repair-journal. pbio. 0040203. g001 "Av Jill O. Fuss, Priscilla K. Cooper - (CC BY 2. 5) via Commons Wikimedia
2. "DNA mismatch repair Ecoli" Av Kenji Fukui - (CC BY 4. 0) via Commons Wikimedia
